This is the story of a plant and a parasitic fungus. The plant is a species of Boechera – rockcress – in the Brassicaceae, i.e. the mustard family. The fungus is one that causes a disease called “rust”, Puccinia monoica.
This is going to get weird and complicated, and may depend on you at least pretending to remember some of your high school biology, so to be easy on us all, I will start with the plant.
Boechera is a genus that includes a couple dozen species, most of which were formerly in the genus Arabis. Like many other small brassicas, rockcress’s main concern in choosing a habitat is to find a place where it doesn’t get drowned, and where it can get in and out before bigger things overtop it. For example, rocky habitats with shallow soil work well. Also like many brassicas, the rockcresses are by and large biennial. They put out a rosette of leaves in the first year, perhaps in late summer or early autumn, then “bolt” in the second year, putting on one to several erect stems with flowers at the tops in a tight and narrow inflorescence. In some cases, these can be as much as 3 feet tall, but generally they are much smaller.
 In the Valley, I have seen this parasite only on what I believe to be B. pauciflora. For now, however, I rather like drawing on the left of one species from the 1912 edition of the Fieldbook of American Wildflowers by F. Schuyler Mathews. It is closely related…
In the Valley, I have seen this parasite only on what I believe to be B. pauciflora. For now, however, I rather like drawing on the left of one species from the 1912 edition of the Fieldbook of American Wildflowers by F. Schuyler Mathews. It is closely related…
Although B. pauciflora itself doesn’t have any special ecological role, plant molecular biologists are interested in boecheras partly because of their small genomes. B. stricta, for example, one of the species found in Idaho, has a genome of about 227 Mbases (that’s considered small) on 7 chromosomes (also small). It is one of two Boechera species whose genomes have been completely sequenced and it has been the subject of several hundred scientific articles… not bad for a small, ephemeral, nondescript, unimportant plant. There is an incredible paucity of information of any sort available on B. pauciflora.
Another very interesting feature of many species in this genus is that they reproduce asexually, producing seeds by apomixis… similar to the way dandelions make their seeds. Nevertheless, they do make pollen, and do attract insects to carry that pollen to other flowers. This is an important part of our story.
On the fungal side, Puccinia is one of about 168 genera of rust fungi; half of all the 7000 rust fungus species are in this genus, including our star, P. monoica. More broadly, all of these are in the larger group, the Basidiomycota, which also includes almost all the fungi (mushrooms) we intentionally eat.
All the rust fungi are obligate parasites, and they have by far the most complex life cycles of any fungi, probably of any eukaryote.
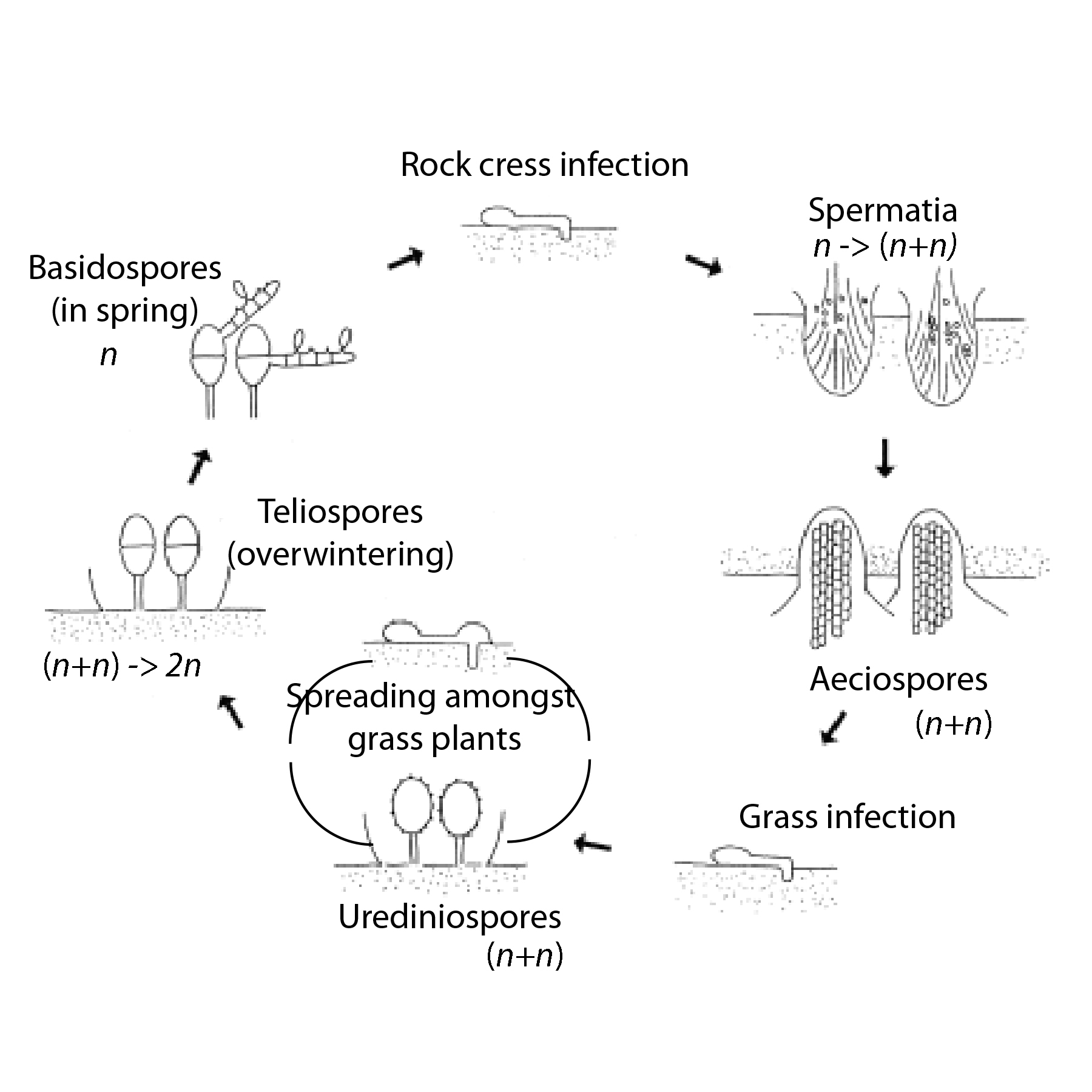
The general life cycle of P. monoica is illustrated in the diagram to the right. It requires two unrelated plants, five stages, and five different kinds of spores, each with its distinct spore producing fruiting bodies. None of these is big enough to qualify as a “mushroom”. Of course, in biology, nothing is simple (not that a 5 part life cycle could be simple), and the rusts can be parasitized by other fungi and their spores eaten a number of species of insects (e.g. gall midges).
Being a basidiomycete, one of the kinds of spores Puccinia produces is a basidiospore. It only produces these on one of its hosts, and by convention, this is called the “primary host”. In this case it is a wild grass of some (unidentified) sort. This stage is a good one to consider the “beginning” of the life cycle, keeping in mind that cycles don’t actually have beginnings any more than bike wheels have ends. This works because the basidiospores are produced in the spring when rock cress growth is starting up again. The critical step is for the basidiospore to land on the Boechera and germinate. The funny little drawing at the top of the illustration depicts a spore (on the left) with a flattened “appresorium” growing out of it and an infection peg (on the right) penetrating the rock cress epidermis.
We’ll get back to how basidiospores are formed, but for now, one important bit of Puccinia genetics is that each has only a single copy of the fungal genome, i.e. it is haploid. We call this n. Our own personal cells, other than eggs and sperm, are diploid… they have two versions of each chromosome and are called 2n. Basidiospores also only have one nucleus so they are monokaryotic.
After the spore has germinated and the mycelium has penetrated into the leaf, it simply goes on growing and spreading, sucking up nutrients from the plant as it goes. That’s what parasites do.
Before finishing the rest of the fungal life cycle, let’s talk first about what else the spreading fungus does. To me as a plant biologist, these are especially cool bits of biology.
As they grow, besides sucking up all the nutrients, the mycelia massively alter gene expression in the plant. The result is that eventually the fungus neuters its host and stops it from forming flowers… and from ever producing seeds. In the language of population ecologists or evolutionary biologists, this makes the fitness of these individuals zero. Instead, the fungus forces our little plant to make what are known as “pseudoflowers.” These are the yellow parts that fool you, at first, into thinking they have petals. Pseudoflowers are basically altered leaves that look and smell like real flowers. In fact, they look like real flowers in both the visible and ultraviolet wavelengths that insect pollinators see. Pseudoflowers also produce insect-attracting nectar-like substances.

Also on each pseudoflower are hundreds of small cups (spermatogonia) that produce spermatia, a second kind of spore. Visiting insects get covered with these and carry them to other infected plants, to other spermatogonia. The bumps on the tops of the pseudoflower “petals” in the photos are the spermatogonia. They are also on the stems, but not on the bottoms of the “petals”.
Individual spermatia are neither “male” nor “female” but of compatible or incompatible “mating types”. At this point, they are also still monokaryotic and haploid. If, after dispersal by the insects, two compatible spermatia contact each other, they fuse, but the nuclei stay separate. This odd behavior means that the resulting mycelium is dikaryotic – each cell contains two dissimilar, haploid nuclei (called n+n, not 2n). Stick with me; this is cool.
Thus infected, next, the pseudoflowers give up the whole charade of producing pigments and the dikaryotic mycelia start into producing the next set of fruiting bodies and their spores. The structures are called “aecia” and the spores, aeciaspores. Often, to avoid crowding or competition, the aecia form on the opposite side of the “petals” and leaves from the spermatogonia. As I write this in mid-April in Teton Valley, aecia seem not yet to have been formed.
Aeciaspores are also dikaryotic (n+n) but everything about their gene expression for structure and function is different from the spermatia; they are configured specifically to carry the infection away from the rock cress to the primary host. In the Valley, they have to wait for this until the grasses start growing again which is some time after the rock cress. Aeciaspores are produced in huge numbers. As they are dispersed by the wind, this at least increases their chance of some finding a suitable host rather than blowing away and rotting in oblivion somewhere.
At this point, I won’t go into details about the grass host; most sources don’t actually suggest what it is other than “some species of grass”. But it is important to finish off this life cycle problem. As shown in the illustration above, the infection process for the grass again involves spore germination, appresorial growth, an infection peg and the gradual takeover of the leaves. It does not, however, involve more pseudoflowers. Instead, on the primary host, as it grows and steals nutrients, the fungus forms yet another spore-producing body, the uredinia which produces urediniospores. These are still dikarytotic (n+n). They are also capable of infecting other grass plants and do so as long as there are plants to infect. It is these structures that give the leaves the rusty color responsible for the name of the fungus and the disease it causes.
But, alas, all good things do come to an end, even for parasitic fungi. Autumn comes, the weather turns cold, the grasses prepare to die back to their roots and rhizomes, and the fungus knows when to call it quits. At that point, the uredinia legally change their names to telia, and start producing teliospores. Teliospores are different in that they can not infect any host, but have thick walls and are quite good at surviving the winter on the dead grass leaves. The dikaryotic cells give up their own charade and the nuclei fuse. We finally have something that is diploid (2n) and monokaryotic. And when spring comes, each of these germinates, producing a basidium. There, meiosis happens, four genetically different basidiospores form (assuring diversity and adaptability of the fungus), and the whole cycle continues.
The Puccinia/Boechera symbiosis is, IMHO, one of the more fascinating in local plants whose manifestation is visible. But I’m not sure if I would put it among the most fascinating of all parasitic arrangements. If, in some macabre way, you like this stuff, consider reading Parasite Rex by Carl Zimmer.
Sources and acknowledgements:
First an acknowledgement: Special thanks to the iNaturalist community, in particular A. J. Wright and Bonnie Semmling who provided the identification of this “flower” as a fungus. A. J. Wright also identified the plant host.
Thomas Roehl, 2016, Order Pucciniales, the Rust Fungi, Fungus Fact Friday #130 … a fascinating set of fungal-centered articles, put out weekly online.
Cano, Liliana M et al. “Major transcriptome reprogramming underlies floral mimicry induced by the rust fungus Puccinia monoica in Boechera stricta.” PloS one vol. 8,9 e75293. 17 Sep. 2013, doi:10.1371/journal.pone.0075293 … a full blown scientific journal article, but bits of the abstract are generally understandable.
Nicholas P. Money, 2016, Fungal Diversity, in The Fungi (Third Edition). The life cycle diagram was modified from here.
Carl Zimmer, 2000, Parasite Rex: Inside the Bizarre World of Nature’s Most Dangerous Creatures.