As a genus, Chara is cool. The plants look sort of like the flowering plants we celebrate generally on this site. But they are algae. Unlike the plants that are mostly emphasized on this site, Chara are also special because single “internodal” cells (stem lookalikes) can be huge. In Fig. 1 here, the single marked cell is about 7 cm long and 1 mm across! “Normal” plant cells are measured in micrometers.
So, what are these plants? Are they rare? Where did they come from?
Chara spp. are found world-wide, at least between 69˚N and 43˚S. But they are environmentally demanding, needing very clean water, preferably with an above neutral pH. Interestingly, you may find either Chara or mosquito larvae in pond or stream water, but not both. Thus, the Teton River is a pretty good place for them. On the other hand, the main image in the gallery here for the species Chara contraria shows plants in a stock tank on a bike trail in Henderson Canyon.
OK, so I’ve identified the local species, but that doesn’t mean I know why. The identification of the genus to species is very difficult… depending on very small morphological features which are themselves quite variable. These days, charaphytologists (and yes, they actually call themselves that) are more likely to rely on DNA bar coding based on the matK gene–from chloroplasts and other plastids–to separate or group the species.
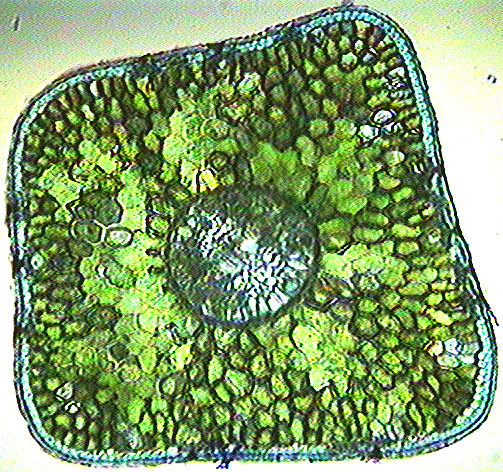
Regardless, all Chara species are multicellular algae with slender green or grey stems. Unlike familiar flowering plants, they don’t have apical meristems from which they grow. They just grow by cell division and specialization all over the plant. The grey color is cool because it results from lime (alkali) incrustations on the outer cell walls. These mask the green color of the chloroplasts. They are also rough feeling, hence the common name, stoneworts. Fig. 2 shows some details of this phenomenon, including that Chara actively raises the pH of the surrounding medium at specific locations on the cell surfaces. (It seems you can also keep Chara on agar, a sort of jello-like medium used widely in plant biology.)
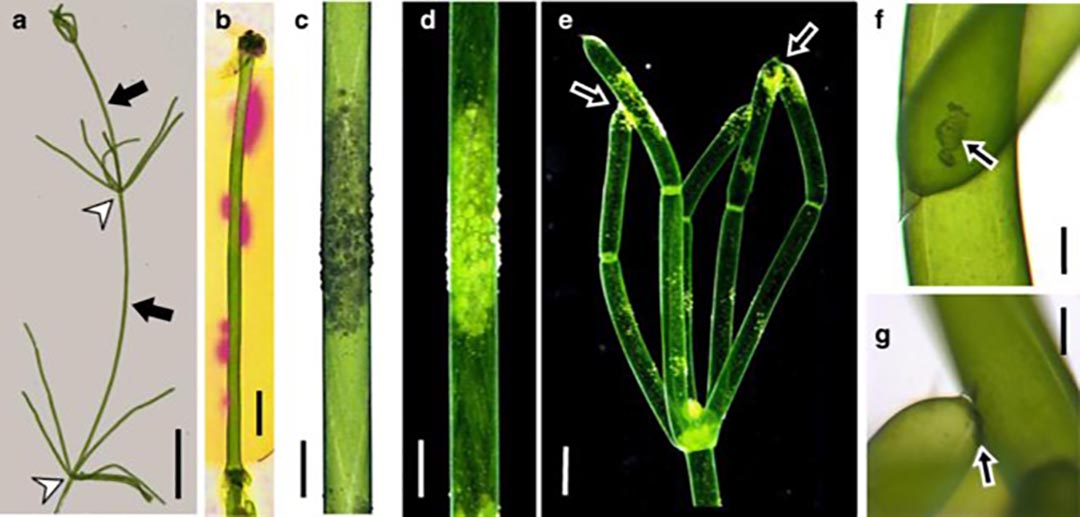
Getting back to the growth form of the alga, well before my time (i.e. in 1623), Characean algae were described as species of Equisetum owing to their superficial resemblance to horsetails (e.g. Equisetum arvense). Although they are not closely related at all, there is an evolutionary connection (see below). So at least superficially, the stoneworts resemble “land plants” in that they have stem-like and leaf-like structures. They also have something resembling roots… rhizoids.
In nearly all Chara species, the internodal cell form the main axis. Often these are reinforced and stabilized by a layer of smaller cortical cells. Certainly, our local species, as shown (sort of) in Fig. 3, has these.

But this is not universal, and in particular, it is not true of Chara australis, Chara braunii, and Chara corallina (Fig. 1).
OK, so what? Is there anything worthy of attention besides this being a sort-of-unusual component of the local ecosystem?
Well, yes, there are a whole slew of reasons. For example, Chara has been an important organism for studying the evolution of what, for lack of a better word, we call “land plants”, or “embryophytes“, i.e. the plants most of interest to those looking at this website. Molecular studies of plant evolutionary relationships, “phylogeny“, have strongly suggested a close relationship between Characeans and those plants… perhaps not to the extent historically suggested that Characeans were the direct ancestor of “land plants”, but close enough. They share, for example, a number of features–the fine structure and pigment composition of chloroplasts, and the composition and structure of their cell walls. (Cell walls are much more than cellulose, and even that isn’t found in all algae.)
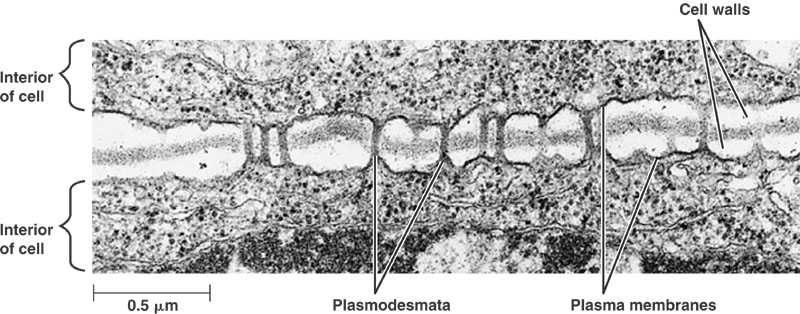
In addition, both groups have cell-to-cell connections through their cell walls, called “plasmodesmata” (Fig. 4). They share a unique mode of cell division, forming “cell plates” between the daughter cells. And they have similar growth controlling substances, a.k.a. hormones, not found in most algae.
For more on the origin of land plants, go here.
Well, and another reason to care… Partly because of this close evolutionary relationship, these species have received a lot of attention by physiologists especially with respect to the transport of ions (nutrient) across membranes and various other membrane processes such as turgor, the pressure that keeps plants rigid (and is that is lost when plants wilt). Those membrane phenomena were what first got me started on liking Chara in the first place. I suppose I got sort of carried away and actually named my cat Chara.
The size of a Chara corallina (or C. australis) internodal cell, for example, and the fact that there are no “interfering” cortical cells around it, make It is easy to impale it with micro-electrodes. Those are very fine-tipped glass pipettes (tubes) that serve as electrical probes to study the voltage, currents, or even ion concentrations across cell membranes. Similar pipettes can be used to “perfuse” these cells, i.e. to pump fluids through them, completely replacing the cytosol and vacuolar contents. This is a handy manipulation if you want to see how transport varies with gradients across membranes. The National Library of Medicine currently lists 800 scientific articles about Chara corallina, many of which were based on just these techniques.
As a teaching tool, Chara corallina is also cool because it is easy to grow, and for demonstrations of phenomena ranging from action potentials (like nerves have, but slower) to growth, to the beautiful phenomenon of cytoplasmic streaming (Fig. 5).
Finally, there’s the question of sex and reproduction. Sorry, but this could get confusing with all the specialized words. You can skip it if you like.
Like other algae, the main plant body of Chara is haploid. That means, unlike you and all the flowering plants we know, the cells have only one set of chromosomes. It’s called a gametophyte, because it can produce gametes (male and female cells). That’s fine if you want to reproduce only by cloning, but really, you want something like a “seed” for better dispersal and adaptation to different environments over time.

And indeed, Characean reproduction does involve sex. The male gametes are produced in a sex organ called a globule (Fig. 6). The females are produced in nucules. In some species (including our local one), the two gametes organs are on the same plants; In that case, the males always develop first. In others species, they are on different plants.
Sexual reproduction results in a zygote that develops into an oospore. This isn’t a seed, per se but it serves the same purpose. The developing oospore, the only diploid stage of the life cycle, has an extremely hard, calcified cell wall which allows survival in wet or dry sediment for many years or even decades. Once good conditions have returned, the oospore cells undergo meiosis (cell division reestablishing the haploid state) as the plant grows into the easily-visible gametophyte.
A bonus feature – Back in the early 1980s, at a meetings in Toronto centered on plant membrane transport, a senior researcher from Adelaide, Australia – F. Andrew Smith– wrote new words to a popular song. To fully understand it, however, you need to be familiar with that song… All around my hat … first recorded in 1976 by Steeleye Span.
In any case, here are the revised words:
All around the cell
There is a band of alkali
All around the cell
And it’s formed in the light
If any one should ask me
Just what it is that forms it
It’s formed by photosynthesis
And it goes away at night
Be well, all